摘要:从中华蜜蜂(Apis.ceranacerana)线粒体DNA中扩增出完整的细胞色素氧化酶II(Ac-COII)和III(Ac-CoIII)基因。两者长度分别为681bp和780bp,Ac-CoII可编码226个氨基酸残基的多肽,与意大利蜜蜂在核苷酸水平和氨基酸水平的一致性分别为90%和95%。Ac-CoIII可编码259个残基的蛋白,与意大利蜜蜂在核苷酸水平和氨基酸水平的一致性分别为84%和79%。利用COII的序列信息进行分析能够反映各不同蜜蜂种类之间的系统进化关系。
关键词:细胞色素氧化酶II和III;中华蜜蜂;序列分析;分子进化
线粒体是细胞重要的细胞器,上面分步许多与呼吸代谢相关的酶类,是生物能量代谢的主要场所,在维系细胞正常功能方面具有重要的作用[1]。线粒体具有自主独立的基因复制和遗传方式,在生物体体内是母系遗传的。因而在研究物种的系统进化关系方面具有重要的意义。围绕不同生物种类的线粒体基因、基因组序列已经完成了许多工作[2-4],对于我们理解线粒体的功能和相关物种的系统进化关系具有重要作用。
意大利蜜蜂的线粒体基因组测序工作已经完成[5],上面总共编码22个转移RNA基因,13个与呼吸代谢密切相关的代谢酶类和2个核糖体基因。由于线粒体基因是母系遗传的,在研究物种之间的系统进化关系具有重要意义,蜜蜂线粒体的许多蛋白编码基因包括NADH,16sRNA,28sRNA,COII,COI,ATPase[6-8]等均曾被用于分析物种之间的系统进化关系。
中华蜜蜂是我国的一个有重要经济价值的蜜蜂种类,在我国大面积饲养。其传统的分类学地位只是在形态学的指标上予以说明。在基因水平上是什么情况还没有工作涉及。本研究克隆了中华蜜蜂细胞色素氧化酶的II单位和III单位基因,并与GenBank内的蜜蜂同源序列进行比对和系统进行关系的分析,以期从分子的角度对我国中华蜜蜂的分类地位予以确定,为相关的研究提供理论基础。
1材料和方法
1.1材料
中华蜜蜂采自浙江大学昆虫研究所蜂场,PGEM–Teasy载体、TaqDNA聚合酶购自Promega公司,Trizol、EcoRI,DNAMarker,低熔点凝胶购自Takara公司,TG1为宿主菌。
1.2方法
1.2.1蜜蜂线粒体基因组DNA的提取
蜜蜂线粒体基因组DNA的提取按照Crozier等[9]的方法进行。大约9g的蜜蜂胸部组织匀浆于300ml0.1MTris-HCL缓冲液,pH7.5,内含0.25M蔗糖,0.01MNaCL,和0.05MEDTA。通过差速离心来收集线粒体,重悬于12mlpH8.0,0.2MTris-HCL缓冲液,内含0.1MEDTA和1%SDS。加蛋白酶K至终浓度为90μg/ml,然后60℃孵育2hr,16000g离心10min,去上清,加2ml5M醋酸钾pH6,冰上放置30min,12000g离心10min,上清经酚、氯仿多次抽提去蛋白,最后酒精沉淀线粒体DNA,溶于适量体积的TE缓冲液,-20℃保存备用。
1.2.2引物设计和目标片段的PCR扩增
用于扩增中华蜜蜂COII和COIII的引物序列参照已经完成的意大利蜜蜂的线粒体基因组序列来进行(GenebankAcession:L06178),在COII和COIII两侧分别设计特异性引物,其中用于扩增COII的引物序列为:F1:5’-ATGGCAGAATAAGTGCAT-3’,R1:5’-CAACTCTTATAATTCAGACATC-3’,用于扩增COIII的引物序列为:F2:5'-CAGCAAATTTAATTTCTGGAC-3',R2:5'-GTAATTGGATTAAATCCACATTC-3'。以蜜蜂的线粒体基因组DNA为模板PCR扩增出目的基因。PCR扩增的条件为94℃变性3min,然后为94℃变性45s,54℃45s,72℃60min,共30个循环。RT-PCR产物用2%琼脂糖凝胶电泳鉴定。低熔点凝胶回收分离目的片段,与载体的连接、转化,酶切鉴定重组克隆。由上海生工基因公司测定DNA序列。
1.2.3序列分析和用于比对的序列来源
测序的结果用GenBank/EMBL内的Blast软件进行蜜蜂同源序列的搜寻和分析,利用CLUSTAL1.8进行序列的多重比对。进化分析利用PAUP(version4.0Beta)软件进行,方法为最大简约法。用于分析的序列来源为(GenBank的接入号):Apismellifera(11990602);Apisdorsata(11761415);Apiskoschevnikovihaplotype1—2(11761411,11761413);Apisceranahaplotype1—8(11761395,11761397,11761399,11761401,11761403,11761405,11761407,11761409);Apisnuluensis(11761393),我们自己的CoII测序结果名为Achz;COIII来自Apismellifera(L06178)。
2结果
2.1C0II和CoIII基因的PCR扩增及基因克隆
以中华蜜蜂的线粒体DNA为模板,设计的特异性引物顺利扩增了编码Ac-COII和Ac-COIII的线粒体DNA片段,大小分别为960bp和1300bp(图1),与预期的大小相符。扩增的片段与PGEM-Teazy载体连接,经酶切鉴定与PCR扩增的大小一致(图2)。
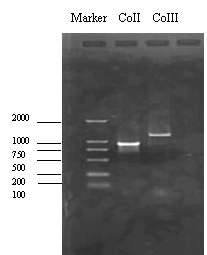
图1从线粒体基因组DNAPCR扩增的COII和COIII
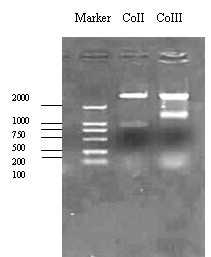
图2扩增的COII和COIII与T载体连接后的酶切鉴定
2.2序列分析
PCR成功扩增了华蜜蜂的完整的COII序列(Ac-CoII)和COIII序列(Ac-CoIII)。其中编码Ac-COII的序列全长为681bp,编码一个226个氨基酸残基的细胞色素氧化酶II单位(图3),在核苷酸水平上,中华蜜蜂Ac-CoII序列较意大利蜜蜂的多三个核苷酸,一致性为90%。Ac-CoIII序列的长度为780bp,可编码一长度为259个氨基酸残基的蛋白,核苷酸水平的一致性为84%(图4)。
一般生物体的线粒体DNAAT含量比较高。在中华蜜蜂的COII里含量为80.7%,而在意大利蜜蜂线粒体内是78.4%。两者均低于意大利蜜蜂线粒体的平均AT%(83%)。在COIII基因内也有相同的趋势,在中华蜜蜂和意大利蜜蜂体内分别为80%和83%。意大利蜜蜂COIII的AT%与线粒体的平均值相同。
1ATTTCTACATGATTCATATTTATGTTTCAAGAATCAAACTCATATTATGCTGATAATTTG60
ISTWFMFMFQESNSYYADNL
61ATTTCATTTCATAATATAGTAATAATAATTATTATTATAATTTCTACTTTAACAGTATAT120
ISFHNMVMMIIIMISTLTVY
121ATTATTATAGATCTTTTTTTAAATAAATTTTCAAATCTATTTTTACTAAAAAATCACAAT180
IIMDLFLNKFSNLFLLKNHN
181ATTGAAATTATTTGAACAGTAATCCCAATTATTATTTTATTAATTATTTGTTTTCCATCA240
IEIIWTVIPIIILLIICFPS
241TTAAAAATTTTATATTTAATTGATGAAATTGTAAATCCATTTTTTTCTGTAAAATCAATT300
LKILYLIDEIVNPFFSVKSI
301GGCCATCAATGATATTGATCCTATGAATACCCTGAATTTAACAATATTGAATTTGATTCA360
GHQWYWSYEYPEFNNIEFDS
361TATATATTAAATTATAGAAATCTAAATCAATTTCGATTACTAGAAACTGATAATCGTATA420
YMLNYSNLNQFRLLETDNRM
421ATTATTCCTATAAATATCCCATTACGATTAATTACAACTTCCACAGATGTAATTCATTCA480
IIPMNIPLRLITTSTDVIHS
481TGAACTGTCCCATCACTTGGGATTAAAGTTGATGCTGTTCCAGGACGAATTAATCAATTA540
WTVPSLGIKVDAVPGRINQL
541AATTTAATTAGAAAACGACCTGGAATCTTTTTTGGTCAATGTTCAGAAATTTGTGGAATA600
NLISKRPGIFFGQCSEICGM
601AATCATAGATTTATACCAATCATAGTAGAATCTACATCATTTAAATATTTTCTTAATTGA660
NHSFMPIMVESTSFKYFLNW
661GTAAACAAACAAAATAATTAA681
VNKQNN*
图3细胞色素氧化酶II单位的核苷酸序列及其编码的氨基酸序列
在所扩增的Ac-COII片段上游同时包含了tRNAleu和随后的一小段非编码序列,该段非编码序列在中华蜜蜂体内是88bp,在意大利蜜蜂体内是201bp。Ac-CoII的下游序列则包含了tRNAArg的基因序列和随后的6bp非编码序列,其在意大利蜜蜂体内为7bp。
扩增的Ac-CoIII序列上游包含部分的ATpase6基因和随后的18bp的非编码序列,该序列在意大利蜜蜂体内是21bp。Ac-CoIII的下游序列则包括一个56bp的非编码区(在意大利蜜蜂体内为70bp)、tRNAGly基因和部分的NADH3基因。
两种蜜蜂的COII在氨基酸水平上有95%的一致性,总计有12个氨基酸残基不同。其中大多数不同的氨基酸残基具有相似的化学性质,只有2个氨基酸残基发生化学性质不同的替代,这种情况对于维系CoII的活性是非常重要的。利用软件对两种蜜蜂的CoII进行疏水性的分析表明两者具有基本一致的疏水性特征。Ac-CoII比意大利的CoII在C末端多出一个氨基酸残基,对酶的活性影响不大。
CoIII在两种蜜蜂之间具有79%的一致性,有33个氨基酸残基发生了变化,其保守性较CoII小许多。可能意味着COIII在功能上较COII的作用要小一些。
1ATGAAAAAAAATTTTCCATTCCATATAGTAACAAATAGACCTTGACCAATCATTTTATCA60
MKKNFPFHMVTNSPWPIILS
61TTTAGGTTAATAAATTCATTAATTGGAACAGCAATCTGAATTTACAGATCAAATATTTTA120
FSLMNSLIGTAIWIYSSNIL
121TTAATAATTCTTAATATAACTAATACAGTAGTAATTATAATATTTTGATTCCGAGATATT180
LMILNMTNTVVIMMFWFRDI
181ATTCGAGAAAGAACATTTCAAGGATTACATACACTTTATGTTATTAATTTCTTAAAATTT240
IRESTFQGLHTLYVINFLKF
241AGTATAGTTCTATTTATTTTATCAGAGCTAATATTTTTTGGTTCTTTTTTTTGAGCCTTT300
SMVLFILSELMFFGSFFWAF
301TTTCATTCATCAGTATCCCCAAATATTGAAATTGATATATTATGACCTCCAAAAGATATT360
FHSSVSPNIEIDMLWPPKDI
361AAATTTTTTGATCCAATAGAAATTCCTTTATTAAATTCATTTATTTTAGTTTCATCCGGG420
KFFDPMEIPLLNSFILVSSG
421TTTGCAATTATTGTTAGACATTATTATATAATTACTAATACTTTAAAATTATCTAAATTA480
FAIIVSHYYMITNTLKLSKL
481TATTTATTTTTAACTATTATATTAGGTACATATTTTATAATTATTCAATTAATTGAATAT540
YLFLTIMLGTYFMIIQLIEY
541GCAAATTCATATTTTTGTTTTAATGATAGAGTTTATGGATCAATTTTTTTTATAGCAACA600
ANSYFCFNDSVYGSIFFMAT
601GGATTTCATGGACTTCATGTATTAATTGGAGTAATTTTCCTTTCTATATCAATTTTACGA660
GFHGLHVLIGVIFLSMSILR
661ATATCAAAAATTCATTTTTCAAATATACATAATATAAATTATGAAATAGCAATTTGATAT720
MSKIHFSNMHNMNYEMAIWY
721TGACATTTTGTAGATGTAATTTGATTATTCTTATATTCATTTATTTATTTATTAATTTAA780
WHFVDVI
WLFLYSFIYLLI*
图4细胞色素氧化酶III单位的核苷酸序列和氨基酸序列
在GenBank内越来越多的核苷酸和蛋白序列使得我们有可能利用CoII序列的信息进行系统进化的分析。我们采用PAUP软件的最大简约法来对已知的蜜蜂CoII序列进行进化分析,结果见图5。我们所测定的中华蜜蜂的CoII序列与其他已经获得的CoII序列很好的落在同一个分支内,其他蜜蜂种类在不同的分支内,能够反应蜜蜂的系统进化关系。
3讨论
克隆的这两个基因所包含的非编码区序列均比意大利蜜蜂的对于序列要小,而位于tRNAleu和COII基因之间的最大的非编码区在东方蜜蜂的体内变异很大,有些地理种群同意大利蜜蜂的相差无几,而另一些则可能完全缺失[10]。因此,从进化的角度上来说,中华蜜蜂更原始一些。
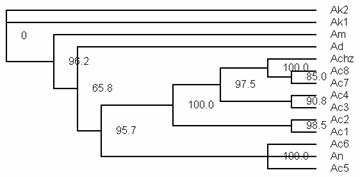
图5基于COII的核苷酸序列对不同种类及倍型的蜜蜂进行的系统进化树的分析结果
从两个亚基的氨基酸序列分析来看,CoII发生改变的氨基酸残基少一些,一致性更高一些,在改变的氨基酸残基里大多是趋于相同性质的氨基酸残基的,对于整个蛋白的结构和功能的影响不大。而CoIII则无论是在一致性、发生改变的氨基酸残基的比例和性质均有较大的变化,因而在蛋白的结构和功能上的变化较CoII要大一些。这种情况表明CoII在进化的过程中更保守,担负的生物功能更重要。相对来说CoIII的重要性要小一些。
蜜蜂的线粒体基因是母系遗传的,在研究分析不同种类之间的系统进化关系时具有重要的意义。迄今为止,已有多种线粒体基因被用于系统进化关系的分析,包括NADH,16sRNA,28sRNA,COII,COI,ATPase等等。随着这方面资料的不断的积累,可以利用基因的序列信息对它们的系统关系进行研究。本研究我们利用COII的核苷酸序列信息,对不同蜜蜂种类的系统关系进行研究,结果表明能够明确的将各种类区别开来,即便是在东方蜜蜂的各不同亚种之间,也比较好的进行了分类。该序列信息是完全可以用于衡量物种系统关系的,具有一定的理论意义。
参考文献:
[1]SuarezR.K,StaplesJ.F.,LightonJ.R.,Mathieu-CostelloO.Mitochondrialfunctioninflyinghoneybees(Apismellifera):respiratorychainenzymesandelectronflowfromcomplexIIItooxygen.JExpBiol.2000,5:905-11
[2]黄朝晖王金福.三种蚊虫COII基因的克隆与序列分析。中国寄生虫学与寄生虫病杂志,2001,19(2):90-92
[3]LiuH.,BeckenbachA.EvolutionofthemitochondrialcytochromeoxidaseIIgeneamong10ordersofinsects.MolPhylogenetEvol,1992,1:41–52
[4]Clary,D.O.andWolstenholmeD.RThemitochondrialmoleculeofDrosophilayakuba:nucleotidesequence,geneorganization,andgeneticcode.J.Mol.Evol.1985,22:252~271
[5]Crozier,R.H.,andCrozier,Y.C.ThemitochondrialgenomeofthehoneybeeApismellifera:Completesequenceandgenomeorganization.Genetics.1993,133:97–117
[6]KoulianosS.andSchmid-HempelP.Phylogeneticrelationshipsamongbumblebees(BombusLatreille)inferredfrommitochondrialcytochromebandcytochromeoxidaseIsequences.MolPhylogenetEvol,2000,14(3):335–341
[7]PedersenB.A.phylogeneticanalysisofcuckoobumblebees(PsithyrusLepeletier)andbumblebees(BombusLatreille)inferredfromsequencesofthemitochondrialgenecytochromeoxidaseI.MolPhylogenetEvol,1996,5(2):289-297
[8]SchmitzJ.,MoritzR.F.SocialityandtherateofrDNAsequenceevolutioninwasps(Vespidae)andhoneybees(Apis).JMolEvol.1998,47(5):606-12
[9]Crozier,R.H.,Crozier,Y.C.andMackinley,A.G.(1989).TheCOIandCO-IIregionofthehoneybeemitochondrialDNA:Evidenceofvariationininsects:mitochondrialevolutionaryrates.Mol.Biol.Evol.1989,6(4):399–411
[10]DeLaRuaP,SimonU.E.,TildeA.C.,MoritzR.F.,FuchsS.MtDNAvariationinApisceranapopulationsfromthePhilippines.Heredity.2000,84:124-30湖北天马养蜂场,加我们的微信一起学养蜂。
福建农林大学蜂学学院李江红
浙江大学应用昆虫研究所张传溪